Huge thanks to Jesamine Bartlett – a recent MSc graduate from the University of Sheffield who has been working on Tardigrade research – for providing this introduction to the weird world of water bears…

Whether or not you like microbiology, bugs, or even science, no one can deny the frankly awesome nature of the Tardigrade. Even the tabloid press are fans! Tardigrades are small, inconspicuous invertebrates that live quietly in almost every habitat we know of. If there is water, there will be probably be active Tardigrades. Even where there isn’t water they have a remarkable ability to enter a hibernation state that allows them to survive freezing and drying out. They have been found at both poles, 20,000ft up mountains in the Himalaya to 14,000ft deep on the ocean floor (Everitt, 1981; DeSmet & Van Rompu, 1994; Ramazzotti & Maucci, 1983; Renaud-Mornant, 1982 ). They are as close as residents in your back garden and as far out as visitors to outer space! More on that later…
Known as moss piglets or water bears because of their clawed legs (called lobopods) and lumbering bear-like gait, the Tardigrade is a tiny creature that measures on average 500µm when adult. They are small enough that you can’t see them with the naked eye, but large enough that you can easily extract them from moss and view them with a good hand lens or simple microscope. Similarly to many invertebrates with a hard outer cuticle, they grow by moulting. This process is known as “ecdysis” and is the same process as spiders shedding their skin. Often Tardigrades take advantage of this discarded hard outer shell by laying vulnerable eggs inside them for protection. Tardigrades reproduce sexually, with females either laying the eggs in the moult with the male fertilizing them afterwards, or through internal fertilization and eggs lain afterwards. Tardigrades can also reproduce via “parthenogenesis” – a hermaphroditic “female” fertilizing her own eggs internally. Tardigrades are generally omnivorous, eating plant cells, algae and also other microscopic invertebrates such as rotifers or even other Tardigrades. Having a flexible diet that relies on no single food source helps water bears to exist in such disparate places across the globe. Because of their ability to survive pretty much anywhere, Tardigrades are one of comparatively few multicellular animals to earn the status “extremophile”, a label that is usually attributed to bacteria and archaea that can happily live in extreme environments such as deep sea hydrothermal vents, or trapped deep in glacial ice with little or no oxygen. Since extremophiles live in such rare and inhospitable conditions, they can tell us about the ranges of conditions where life is possible.

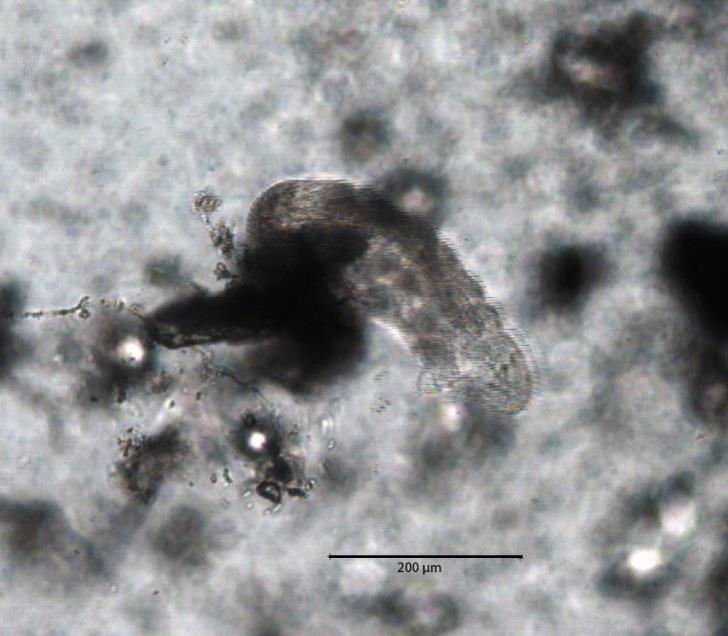
Over the last few decades, the extreme habitats at Earth’s poles have attracted increasing research attention. Complex life has now been identified on, in and beneath Arctic, Antarctic and mountain ice (Wharton et al, 1981; Kohshima 1984; Hodson et al, 2008). Recently, fish were even discovered living beneath lake ice in Antarctica (see also Brent et al 2014), illustrating the potential for previously undiscovered complex life on Earth. At the poles, Tardigrades inhabit deglaciated terrain, and the surfaces of glaciers and ice sheets. Living in and under ice is extreme in the highest degree, with low nutrients, low light, low temperatures and high pressures. But it is comparatively stable to that of the surface; Frequent and drastic environmental fluctuations including periodic freezing and thawing, glacier hydrology, meteorology and intense irradiance challenge incumbent microbes, some of which develop specific physiological adaptations that allow them to survive. In order to survive extremely low temperatures (for example on ice surfaces during winter beneath a seasonal snowpack). Tardigrades enter a state of “cryptobiosis”, slowing their metabolisms to ~0.01% of normal and losing up to 99% of their total moisture. This cold, dry hibernation state is known as their “tun” state. The cessation of metabolism to such levels would kill most animals, but Tardigrades can amazingly reverse this state, blurring the line between living and dead. In the presence of liquid water, such as a spring thaw, Tardigrades can awaken, rehydrate and reanimate themselves. While in the tun state, a Tardigrade can survive for years in adverse environments, and after 10 years most can fully reanimate and continue their life cycle as if they had only just stopped to have a nap. The record for reanimation is 120 years in tun state – although the Tardigrade in question didn’t survive very long after awakening. As a tun, Tardigrades are light, dry capsules of their former selves, capable of being transported about by wind, animals and perhaps even asteroids, which to some extent revived the Panspermia hypothesis – that life on Earth was brought here by a ‘hitchhiking’ organism (Jönsson et al, 2008; Pasini, 2014).
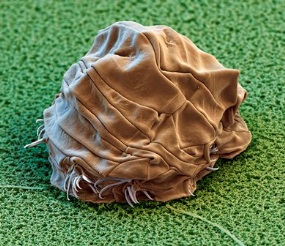
The ability to survive in the extreme cold, even close to absolute zero in some experiments, has led to Tardigrades being studied for their viability in space. In 2007, the European Space Agency (ESA) sent live Tardigrades and unhatched eggs into the vacuum of space beyond the International Space Station, where they orbited the Earth for 12 days, fully exposed to the full spectra of solar radiation. They called the programme “Tardigrades in Space”, or TARDIS for short to the acclaim of many a Dr Who fan! The eggs hatched unharmed and 68% of the adult Tardigrades reanimated with no apparent ill effects. In preparation for a potential launch to Mars in the coming decade through the ExoMars programme, the ESA are currently recreating Martian environments to test the Tardigrades under. So you can be assured that they will continue to be a key to exploring the possibilities of extra-terrestrial life in the future.
One of the keys to a Tardigrade’s ability to reanimate from a tun state and survive such extreme habitats seems to be their ability to turn glucose sugar into trehalose sugar (Teramoto et al 2008). Trehalose is a disaccharide cellular sugar found throughout the natural world that helps prevent molecular desiccation (the process of cell deformation and rupture from drying and/or freezing). Trehalose stores water in a “gel phase” that can form a supportive cast around cell walls and organelles, preventing them from distorting and also reduces the amount of un-bonded water available for freezing. Converting blood sugars into trehalose is not unique to Tardigrades. Bees switch between glucose, trehalose and sucrose depending on their metabolic rates (Blatt & Roces, 2001). Trehalose does not fully explain the resilience of Tardigrades, however. It is still unclear how they withstand drastic fluctuations in solar radiation, pressure or temperature and survive to produce healthy offspring. Understanding these processes might help scientists find ways of better preserving living tissues, whether that is human eggs for fertilization, organ transplant, maybe even one day full human suspended animation!
The physiology of Tardigrades therefore make them extremely interesting invertebrates, but they are also significance players in many ecosystems, in particular the truncated food webs at the poles. And it is in cryoconite holes that Tardigrades are able to thrive on ice. These habitats provide favourable conditions for primary production by forming quasi-stable holes in the ice surface (Cook et al, 2010). Cryoconite holes collect allochthonous organic carbon (Telling et al, 2010), receive nutrients and cells from in-flowing meltwater (Irvine-Fynn and Edwards, 2013) and prevent the redistribution of debris for long enough for a multi-trophic ecosystems to develop (Hodson et al, 2008). And where there is an active ecosystem, carbon is cycled. The carbon potential of glaciers and ice sheets has been estimated to be equivalent to terrestrial soil systems (Anesio et al, 2009), and the cryoconite ecosystem could potentially be a large contributor to that (Cook et al, 2012). My work has begun to explore the role of Tardigrades in the cryoconite hole’s carbon cycle, a process they contribute to by oxidizing carbon stored in organic molecules within the cryoconite micro-habitat by grazing on algae and cyanobacteria. And recent data suggests that they could be a significant component in Svalbard cryoconite, particularly during the autumn months when they potentially contribute to cryoconite holes turning from a carbon sink to a carbon source with the onset of seasonal snowfall.
Although small and innocuous, Tardigrades might play a huge role in developing our understanding of the limits and origins of life. Where they exist, their role in biogeochemical cycling and microbial ecology needs to be better understood, especially in truncated glacial food webs where their top-down controls upon community ecology might be important.
To summarise, I think we should all pay homage to the space-venturing, ice-surviving, reanimating microscopic “hoover-bag”: The water-bear, the moss-piglet, the Tardigrade!
J Bartlett
References:
Anesio, A., Hodson, A., Fritz, A., Psenner, R. & Sattler, B. (2009). High microbial activity on glaciers: importance to the global carbon cycle. Global Change Biology. 15: 955–960.
Blatt, J., & Roces,F. (2001). Haemolymph sugar levels in foraging honeybees (Apis Mellifera carnica): Dependence on metabolic rate and in vivo measurement of maximal rates of trehalose synthesis. Journal of Exp. Bio. 204: 2709–2716.
Cook, J., Hodson, A., Telling, J., Anesio, A., & Bellas, C. (2010). The mass – area relationship within cryoconite holes and its implications for primary production. Journal of Glaciology. 51: 106–110.
Cook, J., Hodson, A., et al. (2012). An improved estimate of microbially mediated carbon fluxes from the Greenland ice sheet. Journal of Glaciology. 58:1098-1108.
De Smet, W., & Van Rompu, E. (1994). Rotifera and Tardigrada from some cryoconite holes on a Spitsbergen (Svalbard) glacier. Belg J Zool. 124: 27–37.
Everitt, D. (1981). An ecological study of an Antarctic freshwater pool with particular reference to Tardigrada and Rotifera. Hydrobiologia. 83: 225-237.
Hodson, A., Anesio, A., Tranter, M., Fountain, A., Osborn, M., Priscu, J., Laybourn-Parry, J., & Sattler, B. (2008). Glacial ecosystems, Ecological Monographs. 78: 41–67.
Irvine-Fynn, T.D.L., Edwards, A., Newton, S., Langford, H., Rassner, S., Telling, J., Anesio, A., Hodson, A.J. (2013). Microbial cell budgets of an Arctic glacier surface quantified using flow cytometry. Environmental Microbiology. 14: 2998 – 3012.
Jönsson, K. I., Rabbow, E., Schill, R. O., Harms-Ringdahl, M. & Rettberg, P. (2008). Tardigrades survive exposure to space in low Earth orbit. Curr. Biol. 18: 729-731.
Kohshima, S., (1984) A novel, cold tolerant insect found in a Himalayan glacier. Nature. 310: 225-227.
Nelson, D.R. (2001). Current status of the Tardigrada: evolution and ecology. Integrative and comparative biology. 42: 652–9.
Pasini, D., & Price, M. (2014) Panspermia survival scenarios for organisms that survice typical hypervelocity solar system impact events. European Planetary Science Congress 2014, EPSC Abstracts, Vol. 9, id. EPSC2014-68.
Brent, C., Priscu, J et al. (2014) A microbial ecosystem beneath the West Antarctic ice sheet. Nature 512:310–313.
Ramazzotti, G., & Maucci, W., (1983). Il Phylum Tardigrada. III edizione riveduta e aggiornata. Mem. Ist. Ital. Idrobiol, 411-1012 [NB: English translation is available from Dr. Clark Beasley, Biology Dept., McMurry University, Abilene, TX, USA 79697].
Renaud-Mornant, J. (1982). Species diversity in marine Tardigrada. In D. R. Nelson (ed.), Proceedings of the third international symposium on the Tardigrada, August 3–6, 1980, Johnson City, Tennessee, pp. 149–177. East Tennessee State University Press, Johnson City.
Telling, J., Anesio, A. M., Hawkings, J., Tranter, M., Wadham, J. L., Hodson, A. J., & Yallop, M. L. (2010). Measuring rates of gross photosynthesis and net community production in cryoconite holes : a comparison of field methods. Annals of Glaciology. 51:153–162.
Teramoto, N., Sachinvala, N., Shibata, M. (2008). Trehalose and trehalose-based polymers for environmentally benign, biocompatible and bioactive materials. Molecules. 13:1773-816.
Wharton, R.A., Vinyard, W.C., Parker, B.C., Simmons, G.M., Seaburg, K.G. (1981) Algae in cryoconite holes on Canada glacier in southern Victoria Land, Antarctica. Phycologia. 20: 208-211

5 thoughts on “Water Bears on Ice: Guest blog by Jesamine Bartlett”