I recently published an article in French pop-sci magazine La Recherche about the wondrous microbial ecosystems on glaciers and ice sheets (here for French speakers). For those English speakers who do not subscribe to la Recherche, here is a translation.
Also, I strongly recommend the excellent translator who worked on this article with me – contact me if you need translation services and I can link you up.
The microbes accelerating glacier melting
Context
Our planet is getting warmer and losing its ice. Mountain glaciers are disappearing and the great Greenland and Antarctic ice sheets are shrinking. These masses of ice are giant coolers for the planet and they reflect energy from the Sun back out into space, meaning the smaller they become, the more the planet warms. Surprisingly, the process of melting the vast glaciers and ice sheets is accelerated by microscopic life.
Glacier and ice sheet melting depends upon more than just temperature. Most of the energy driving melt comes from sunlight that hits the ice surface. Dirtier, darker ice absorbs more solar energy than clean, bright ice meaning more energy is available to drive melting. On the Greenland Ice Sheet in particular, the ice becomes very dark in the summer, with large areas reflecting just 20-30% of the sunlight hitting them. This is not a new phenomenon – in fact it was noticed by explorers during the great polar expeditions of the late 1800s. Intrigued, they examined samples of ice under their microscopes. The dark colour of the ice was not simply due to dust as they expected – astonishingly, the ice was stained by life (Nordenskjold, 1875). The ice surface is a patchwork of greys, reds and purples coloured by the collective effect of countless microscopic organisms, with potential knock-on effects for Earth’s climate (Uetake et al., 2010; Takeuchi et al., 2006; Yallop et al., 2012; Cook et al., 2017).

Microbes on Ice
When explorer Adolf E Nordenskjold arrived on the Greenland Ice Sheet in 1870 he immediately noticed the dark grey-purple colour of the ice. His colleague, a biologist called Berggren, examined the ice under the microscope and discovered a rich variety of microbial life. The importance of their discovery was clear to them – this life darkens the ice and increases its melt rate. Nordenskjold even suggested that the microbial life was the “greatest enemy of the mass of ice” and an accelerator of deglaciation at the global scale (Nordenskjold, 1875)!
Until recently, Nordenskjold’s observations of life on ice have remained obscure footnotes in the history of Polar exploration; however, as climate science has become increasingly urgent in the twenty-first century, Nordenskjold’s work has gained new significance. Contemporary scientists have confirmed the presence of a microbial ecosystem growing on the surface of the Greenland Ice Sheet and elsewhere and are now attempting to quantify their ice-darkening effect. Although it is an extreme environment where temperatures are low and nutrients scarce, there is abundant sunlight and liquid water to support photosynthesis, meaning microalgae can grow on the ice surface (Uetake et al., 2010; Yallop et al., 2012). The days are long in the Arctic in summer, with the sun staying above the horizon for twenty-four hours per day for part of the season, exposing the algae to intense and prolonged solar energy. This powers photosynthesis but over time the exposure stresses the ice algae, causing them to produce biological sunscreen molecules to protect their delicate photosynthetic machinery. These ‘carotenoids’ colour their cells very dark purple and enhance the biological darkening of the ice surface.
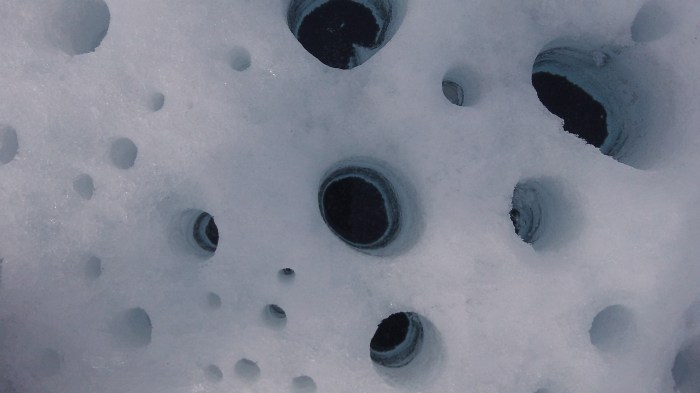
At the same time, the ice surface is peppered with holes that are often cylindrical but can have complex and irregular shapes (Cook et al., 2015). These holes range from centimeters to meters in diameter and depth and contain mixtures of biological and nonbiological material bundled up into small balls that sit on the hole floors. Nordenskjold first noticed these holes on the Greenland Ice Sheet and named them ‘cryoconite holes’, from the Greek for ‘holes with frozen dust’. These holes are the most biodiverse microbial habitat on Earth’s ice. They form when dust and debris becomes tangled up by long, thread-like cyanobacteria. The cyanobacteria are photosynthetic and as they grow they exude polymers that act as biological glues, binding the bundles of material together into stable granules. This biological bundling and binding of material creates a microhabitat for other microbes, especially those that can feed on molecules produced by the photosynthesizing cyanobacteria. As the granules grow they become heavier, meaning they settle on the ice surface. The biological material makes them especially dark, so the ice underneath melts quickly, causing holes to form in the ice surface with the granules sitting on the hole floor. The holes provide protection from the weather and intense sunlight and also prevent the microbes from being washed away. The cyanobacteria therefore sculpt the ice surface and engineer a comfortable, stable habitat where diverse microbial life can thrive in this extreme environment.
Cryoconite holes are more than icy buckets that hold microbial life. They are more like microbial mini-cities on ice, with each connected to many others by meltwater flowing between ice crystals just under the ice surface. Cryoconite microbes engage in engineering and construction, production, consumption, competition, predation, growth, reproduction, death, decay, immigration and emigration. There is both import and export of nutrients, waste and other biological material. At the same time, the hole itself changes its shape and size in response to changing environmental conditions with the emergent effect of maintaining the light intensity at the hole floor, promoting photosynthesis (Cook et al., 2010). Algal blooms and cryoconite are crucial components of the wider Arctic ecosystem, acting as stores of carbon (which they draw down from the atmosphere and fix into organic molecules), nutrients and biomass which can all be delivered to soils, rivers and oceans as glaciers melt (Stibal et al., 2012). Truly, these are widely interconnected complex adaptive systems created biologically on Earth’s ice.
The Cutting Edge of Life on Ice
While life on ice has been known for many years, most of the literature on the subject has been produced during the twenty-first century. Modern molecular biological techniques have enabled scientists to catalogue the species present in cryoconite and algal blooms, and modern instruments can measure their darkening effect. However, there are several major gaps in our understanding of life on ice. To quantify their effect on ice darkening worldwide, we need a reliable method to map icy microbes at the scale of entire ice sheets. From a biological perspective, we know which organisms live in algal blooms and cryoconite so we must now concentrate on determining how they function and what ecosystem services they might provide that could impact human society.
To estimate the total coverage of life on ice, we must detect it without actually being present to take samples. It is relatively easy to take samples and analyse them in a laboratory to tell if life is present, but doing the same from the air is a different problem. In addition to biological darkening, soots and mineral dusts colour the ice. Also, as the ice melts the crystals change shape and melt water can fill the spaces between them, which in itself changes the way the ice absorbs and reflects solar energy. Disentangling the biological signal from these other darkening processes has proven to be challenging.
However, because the darkening of ice by living cells is due to biological molecules that absorb light at specific wavelengths, we may be able to use the spectrum of reflected light to identify them. Chlorophyll, for example, absorbs red and blue light much more effectively than it absorbs green light (which is why we see leaves as green). For other biological molecules, the peak absorption will be at slightly different wavelengths, and non-biological materials will have their own absorption patterns too. However, while identifying ‘signature spectra’ is simple when only one material is present, it is much more difficult when several species with different light absorbing properties are mixed with non-biological materials. All of the light absorbers can be scattered unevenly and mixed vertically within the volume of ice which can itself be a complex aggregate of variously sized ice crystals and liquid water. The reflected light is a tangle of signals that can be hard to unpick.
At our laboratory at the University of Sheffield, we are working on a purpose-built drone which will fly back and forth over a patch of the Greenland Ice Sheet taking images at specific wavelengths of light. By analysing these images we hope to be able to produce a map of life on ice. Using the drone means we can follow the flight on foot and take ground samples to examine in the laboratory, enabling us to link the drone images to actual concentrations of different light absorbers on the ground. The wavelengths imaged by the drone match up with those measured by several Earth observation satellites, meaning that achieving life-detection using a drone should then enable the same from space.

As well as knowing where the life is, we also need a deeper understanding of how it functions. Recognition of ice surfaces as microbial habitats came at the same time as an explosion in accessible and affordable techniques in field molecular microbial ecology, meaning several groups have used high-throughput sequencing of marker-genes to identify the particular microbes present within cryoconite communities (e.g. Cameron et al., 2012; Edwards et al., 2014; Stibal et al., 2014, 2015). Environmental genomic techniques have also been used to investigate the total genetic composition of cryoconite communities (Edwards et al., 2013). To date, these have been snapshot studies, but in the very near future great insights into the functioning of cryoconite microbes will come from rapid metagenomic, metabolomic and metatranscriptomic studies. It has been suggested that ice surface microbes might be good targets for bioprospecting. Since they are able to thrive in conditions of low temperature, high light and low nutrients, they may well utilize survival strategies that we can exploit, either by extracting novel genes and biomolecules, or by observing and gaining ecological knowledge. Cryoconite has been suggested to be a potential source of antifreeze proteins, novel antibiotics and cold-active enzymes. The shape, illumination conditions and flushing with flowing meltwater make cryoconite holes natural analogs to industrial bioreactors which are commonly used to synthesise valuable biomolecules (Cook et al., 2015).
Deep insights will come from combining the expertise of microbial ecologists with glaciologists and physicists who, together, will link processes operating at the molecular level with changes in ice surface colour and patterns of melt, which suggests insights into the ecology of ice surfaces might one day be obtainable from the sky or from space. While this is some way off, great insights could be gained from a shift towards a holistic understanding of the ice surface as a ‘living landscape’.
Extraterrestrial Ice
We are working hard to achieve remote detection of life on ice for the purposes of mapping biological ice darkening from satellites and improving our ability to predict future ice melt. However, there is another potential outcome from this work… what if instead of looking down from space at our own planet, we turn the sensors around and start looking out?
The Greenland Ice Sheet is, in many ways, a good place for developing life detection technologies that can be applied to the search for life on other icy planets and moons. Take, for example, Europa. A recently funded NASA project will examine this icy moon of Jupiter for signs of life because of its potentially habitable icy shell and subsurface ocean. On Europa, the icy surface is sunlit and seeded with possibly mineral-rich snow that forms when liquid water in its subsurface oceans escapes via huge geysers (Hand et al., 2017). There is therefore a potentially dusty ice surface illuminated by sunlight that could support photosynthesis, just like the Greenland Ice Sheet (although the solar energy flux and temperature is lower on Europa and photosynthesis is highly unlikely). Any life detection technology that works on the Greenland Ice Sheet will have to overcome the challenges of ice optics, interference by mineral dusts and uncertain biological pigment composition, which would also be the main challenges for remote detection of life on the surface of other icy planets and moons. The frontiers of glacier biology on Earth may therefore intersect with the cutting edge search for extraterrestrial life.

Summary
While many people think of Arctic and Antarctic ice as lifeless places, there is in fact abundant microbial activity on Earth’s glaciers and ice sheets. But more surprising is the huge impacts of these tiny organisms. By changing the colour of the ice surface, microbes are potentially enhancing the rate at which glaciers and ice sheets are shrinking, but we cannot yet build them into our climate models. The research priority now is mapping these ecosystems from space because this will enable us to estimate their impact on ice melt worldwide and improve our melt forecasts. The same technologies that will enable us to detect life on Earth may eventually be useful tools for searching for icy life elsewhere in the universe. There is also much to be learned about way these microbes function that can educate us about the limits of life in extreme environments. The true sharp edge of glacier biology research involves understanding how these microbes are able to sense, survive and drive environmental change. The study of life on Earth’s ice is deeply interdisciplinary and ultimately it requires us to recognize – as Nordenskjold did – the intricate bridges joining the very big and the very small.
References
Cameron K, Hodson A J, Osborn M (2012) Carbon and nitrogen biogeochemical cycling potentials of supraglacial cryoconite communities. Polar Biology, 35: 1375-1393
Cook J, Hodson A, Telling J, Anesio A, Irvine-Fynn T, Bellas C (2010) The mass-area relationship within cryoconite holes and its implications for primary production. Annals of Glaciology, 51 (56): 106-110
Cook, J.M., Edwards, A., Irvine-Fynn, T.D.I., Takeuchi, N. 2015. Cryoconite: Dark biological secret of the Cryosphere. Progress in Physical Geography, 40 (1): 66 -111, doi: 10.1177/0309133315616574Cook et al., 2017
Edwards A, Pachebat J A, Swain M, Hegarty M, Hodson A, Irvine-Fynn T D L, Rassner S M, Sattler B (2013) A metagenomic snapshot of taxonomic and functional diversity in an alpine glacier cryoconite ecosystem. Environmental Research Letters, 8 (035003): 11pp
Edwards A, Mur L, Girdwood S, Anesio A, Stibal M, Rassner S, Hell K, Pachebat J, Post B, Bussell J, Cameron S, Griffith G, Hodson A (2014) Coupled cryoconite ecosystem structure-function relationships are revealed by comparing bacterial communities in Alpine and Arctic glaciers. FEMS Microbial Ecology, 89 (2): 222-237
Hand, K.P., Murray, A.E., Garvin, J.B., Brinckerhoff, W.B., Christner, B.C., Edgett, K.S., Ehlmann, B.L., German, C.R., Hayes, A.G., Hoehler, T.M., Horst, S.M., Lunine, J.I., Nealson, H.H., Paranicas, C., Schmidt, B.E., Smith, D.E., Rhoden, A.R., Russell, M.J., Templeton, A.S., Willis, P.A., Yingst, R.A., Phillips, C.B., Cable, M.L., Craft, K.L., Hofmann, A.E., Nordheim, T.A., Pappalardo, R.P., and the Project Engineering Team (2017). NASA, Report of the Europa Lander Science Definition team. Posted Feb 2017. https://solarsystem.nasa.gov/docs/Europa_Lander_SDT_Report_2016.pdf
Stibal M, Sabacka M, Zarsky J (2012a) Biological processes on glacier and ice sheet surfaces. Nature 1554 Geoscience, 5: 771-774
Stibal M, Schostag M, Cameron K A, Hansen L H, Chandler D M, Wadham J L, Jacobsen C S (2014) Different 1558 bulk and active microbial communities in cryoconite from the margin and interior of the Greenland ice 1559 sheet. Environmental Microbiology Reports, DOI: 10.1111/1758-2229.12246
Stibal, M., Schostag, M., Cameron, K. A., Hansen, L. H., Chandler, D. M., Wadham, J. L. and Jacobsen, C. S. (2015), Different bulk and active bacterial communities in cryoconite from the margin and interior of the Greenland ice sheet. Environmental Microbiology Reports, 7: 293–300. doi:10.1111/1758-2229.12246
Takeuchi, N., Dial, R., Kohshima, S., Segawa, T., Uetake, J., 2006. Spatial distribution and abundance of red snow algae on 35 the Harding Icefield, Alaska derived from a satellite image. Geophysical Research Letters, 33, L21502, doi:10.1029/2006GL027819
Uetake, J., Naganuma, T., Hebsgaard, M. B., and Kanda, H. 2010. Communities of algae and cyanobacteria on glaciers in west Greenland. Polar Sci. 4, 71–80. doi: 10.1016/j.polar.2010.03.002
Yallop, M.L., Anesio, A.J., Perkins, R.G., Cook, J., Telling, J., Fagan, D., MacFarlane, J., Stibal, M., Barker, G., Bellas, C., 25 Hodson, A., Tranter, M., Wadham, J., Roberts, N.W. 2012. Photophysiology and albedo-changing potential of the ice-algal community on the surface of the Greenland ice sheet, ISME Journal, 6: 2302 – 2313
