The EGU annual meeting in Vienna is one of the major events in the earth science calendar, where the latest ideas are aired and discussed and new collaborations forged. My talk this year was in the “Remote Sensing of the Cryosphere” session. Here’s an overview:
Albedo is a primary driver of snow melt. For clean snow and snow with black carbon, radiative transfer models to an excellent job of simulating albedo, yet there remain aspects of snow albedo that are poorly understood. In particular current models do not take into account algal cells that grow and dramatically discolour ice in some places (except our 1-D BioSNICAR model) and few take into account changes in albedo over space and time.
This led me to wonder about using cellular automata as a mechanism for distributing albedo modelling using radiative transfer over three spatial dimensions and time, and also enabling a degree of stochasticity to be introduced to the modelling (which is certainly present in natural systems).
Cellular automata are models built on a grid composed of individual cells. These individual cells update as the model progresses through time according to some function – usually a function of the values of the neighbouring cells. Cellular automata have been used extensively to study biological and physical systems in the past – for examples Conway’s Game of Life, Lovelock’s DaisyWorld and Bak’s Sandpile Model not only gave insight into particular processes, but arguably changed the way we think about nature at the most fundamental level. Those three models were epoch-changing for the concepts of complexity and chaos theory.

For the snowpack, I developed a model I am calling CASPA -an acronym for Cellular Automaton for SnowPack Albedo. CASPA draws on a cellular automaton approach with a degree of stochasticity to predict changes in snowpack biophysical properties over time
At each timestep the model updates the biomass of each cell. This happens according to a growth model (an initial inoculum doubles in biomass). This biomass has a user-defined probability of growing in situ (darkening that cell) or spreading to a randomly selected adjacent cell. Once this has occurred, the radiative transfer model BioSNICAR is called and used to predict the albedo, and the energy absorbed per vertical layer. The subsurface light field is visualised as the planar intensity per vertical layer, per cell. The energy absorbed per layer is also used to define a temperature gradient which is used to drive a grain evolution model. In the grain evolution model, wet and dry grain growth ca occur, along with melting, percolation and refreezing of interstitial water. This is consistent with the grain evolution model in the Community Land Model. The new grain sizes are fed back into SNICAR ready for the albedo calculation at the next timestep.
At the same time, inorganic impurities can be incorporated into the model. These include dust and soot. These can be constant throughout the model run, or can vary according to a user-defined scavenging or deposition rate. They can also melt-out from beneath, by having the inorganic impurities rising up through successive vertical layers per timestep.

In this way, the albedo of a snowpack can be predicted in three spatial dimensions plus time. Taking the incoming irradiance into account, the radiative forcing can be calculated at each vertical depth at each cell per timestep. Furthermore, the energy available as photosynthetically active radiation in each layer can be quantified. Ultimately. these values can feed back into the growth model. Coupling the CASPA scheme wit a sophisticated ecological model could therefore be quite powerful.
By default the model outputs a 2D albedo map and a plot of biomass against albedo. It is interesting to realise that the subtle probabilistic elements of the cellular model can lead to drastically different outcomes for the biomass and albedo of the snowpack even with identical initial conditions. This is also true of natural systems and the idea that an evolving snowpack can be predicted using a purely deterministic model seems, to me, erroneous. There are interesting observations to make about the spatial ecology of the system. Even this simplified system can runaway into dramatic albedo decline or almost none. It makes me wonder about natural snowpacks and the Greenland dark zone – how much of the interannual variation emerges from internal stochasticity rather than being a deterministic function of meteorology or glaciology?
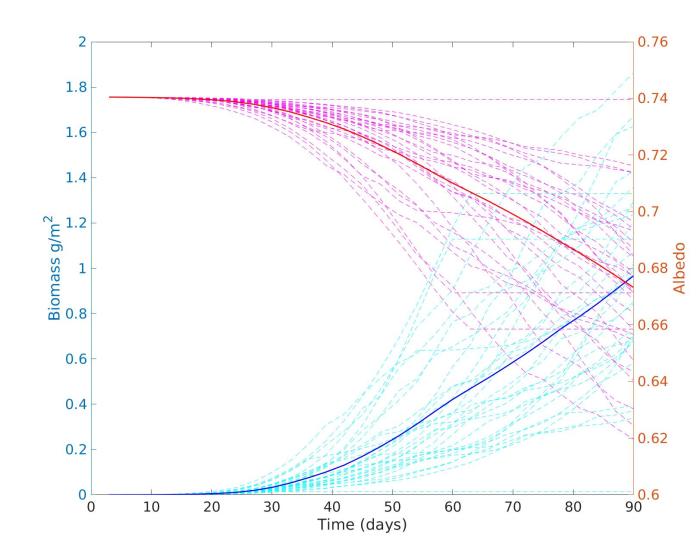
In terms of quantifying biological effects on snow albedo, CASPA can be run with the grain evolution and inorganic impurity scavenging models turned ON or OFF. Comparing the albedo reduction taking into account the physical evolution of the snow with that when the snow physics remain constant provides an estimate of the indirect albedo feedbacks and the direct albedo reduction due to the algal cells.
This modelling approach opens up an interesting opportunity space for remote sensing in the cryosphere. In parallel to this modelling I have been working hard on a supervised classification scheme for identifying various biological and non-biological ice surface types using UAV and satellite remote sensing products. Coupling this scheme with CASPA offers an opportunity to upsample remote sensing imagery in space and time, or to set the initial conditions for CASPA using real aerial data and then experimenting with various future scenarios. At the moment, I lack any UAV data for snow with algal patches to actually implement the workflow, but it is proven using multispectral UAV data from bare ice on the Greenland ice sheet. When I obtain multispectral data for snow with algal blooms, it is possible to automate the entire pipeline from loading the image, classifying it using a supervised classifier, converting it into an n-dimensional array that can be used as an initial state for the CASPA cellular automaton, whose conditions can be tweaked to experiment with various environmental scenarios.
Therefore, the limiting factor for CASPA at the moment is availability of multispectral aerial data and field spectroscopy for training data for algal blooms on snow. In the spirit of open science and to try to stimulate a development community, I have made this code 100% open and annotated despite being currently unpublished, and I’d be delighted to receive some pull requests!
In summary, I suggest coupling radiate transfer with cellular automata and potentially remote sensing imagery is a promising way to push albedo modelling forwards into spatial and temporal variations and an interesting way to build a degree of stochasticity into our albedo forecasting and ecological modelling.
