Nutrient cycling has been a central theme of glacier microbiology in the twenty-first century. Here is a run-down of the fundamentals, focussing on the major ones: nitrogen and carbon. Nitrogen’s up first…
The Nitrogen Cycle:
Nitrogen is a key nutrient required for synthesising crucial organic molecules such as nucleotides, proteins, and chlorophyll. Nitrogen availability also impacts upon rates of primary production (crucially photosynthesis) and respiration in microbial communities.
Nitrogen makes up the majority of earth’s atmosphere, but in its atmospheric form (dinitrogen, N2) organisms cannot use it. It has to be converted, or ‘fixed’ before it can be used to build organic molecules. Just as vinyl records cannot be played on an mp3 player – the analogue information has to be digitised and formatted first – so it is with nitrogen! Organisms cannot use gaseous N2, but when it is fixed into ammonia (NH3) it becomes ‘formatted’ for biota. Once it is ‘bio-available’, it can go through various permutations for incorporation into different biological pathways. Being a cyclic process, organic nitrogen is also ultimately converted back into atmospheric N2.
Nitrogen fixation (N2 -> NH3) requires lots of energy due to dinitrogen’s triple bond and the few organisms (prokaryotes) able to do it do so via complex biochemical mechanisms, sometimes involving host organisms. Nitrogen fixers are diverse but they all use an enzyme called nitrogenase to fix nitrogen. Humans have also learned to force dinitrogen to react with hydrogen to form ammonia through the Haber-Bosch process and release of various bio-available nitrogenous compounds also occurs during combustion of fossil fuels.

For plants to take-up nitrogen through their roots, ammonia must be converted into nitrate (NO3–), primarily via the action of nitrifying bacteria (NH3 -> NO2– -> NO3–). Nitrates are either converted directly back to atmospheric dinitrogen by denitrifying bacteria, or taken up by plants and assimilated into a range of crucial biomolecules. Ultimately, the plants die and decompose, and the assimilated nitrogen undergoes ammonification (back to NH3) and then nitrification (back to NO3–).
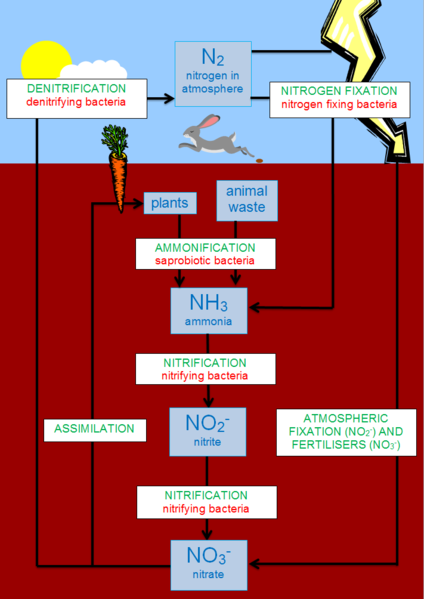
In oceans and lakes the cycle is essentially the same, except that some of the sources and transfers are modified to reflect the aquatic environment. This probably better reflects nitrogen cycling in most supraglacial habitats. Cyanobacteria fixes most of the N2 which enters the system via runoff, precipitation, snowmelt and atmospheric exchange. Bacteria convert ammonia (NH3) into ammonium (NH4+) which can then be nitrified into nitrite (NO2–) and nitrate (NO3–) for assimilation into organic molecules.
Nitrogen Cycling on Ice Surfaces:
Nitrogenous material on ice was first identified by Bayley (1891), but N cycling in the supraglacial environment was largely overlooked until relatively recently. N cycling was examined in polar rivers (Tockner et al, 2002), oceans (Dittmar, 2001), soils (Nordin et al, 2004), ice cores (Olivier et al, 2006), subglacial sediments (Wynn et al, 2007) and under seasonal snow (Williams et al, 1996; Hodson et al, 2005; Hodson et al, 2006; Jones, 1999), but organisms directly inhabiting ice surfaces were not considered in detail until very recently.
Tranter et al (2004) examined Antarctic cryoconite hole biogeochemistry, reporting high inorganic : organic N ratios. This suggests recycling of organic compounds has dominated over fixation of N2, probably due to surface freezing isolating holes from nitrogen inputs. In contrast, Hodson et al (2008) explained that Svalbard cryoconite holes generally show net NH3 production, suggestive of fixation of atmospheric N2 by cyanobacteria. Since nitrogen fixation is so energy demanding, microbes use allochthonous (delivered from elsewhere) bio-available nitrogen in preference to fixing it. Therefore, active fixation indicates that allochthonous bio-available nitrogen is not meeting microbial demand. Sawstrom (2009) confirmed that Svalbard cryoconite communities were not limited by nitrogen availability, highlighting the action of nitrogen-fixing bacteria as important components of microbial assemblages.
In 2009 and 2010, Jon Telling and others undertook detailed investigations into nitrogen cycling across the ablation zone of glaciers in Svalbard and on the Greenland ice sheet. Nitrogen fixation was found to be active in cryoconite hole communities in both regions, implying that there is not enough bio-available nitrogen delivered to these habitats via abiotic processes (predominantly wind and snowmelt inputs) to sustain microbial growth. As mentioned earlier, nitrogen fixation requires a large amount of energy and may therefore exert some limits on cell proliferation.
Some evidence of denitrification and ammonification was also identified in Greenland cryoconite holes by Telling et al (2011), although not quantified. These processes are crucial to nitrogen cycling and are expected to occur in these habitats.

Human Impacts on Glacial Nitrogen Cycling
Greater allochthonous (delivered from elsewhere) nitrogen deposition unburdens microbial nitrogen fixers from the task of providing bio-available nitrogen, meaning more energy is available for microbial growth and proliferation in the community (although nitrogen limitation is unlikely to be the primary factor limiting biomass production in cryoconite holes; Telling et al, 2011). Human derived nitrogenous compounds such as those produced by fossil fuel combustion are being deposited in polar snowpacks in ever increasing concentrations such that Arctic cryoconite communities often no longer need to fix atmospheric nitrogen at all; microbial activity can be sustained by allochthonous inputs (Telling et al, 2011). Hodson et al (2010) monitored nutrient budgets in Svalbard after a particularly intense period of pollutant deposition in Svalbard and showed that even isolated deposition events can have huge impacts for nutrient cycling which permeate through entire glacial catchments.
Anthropogenic activities are increasing the concentration of bio-available nitrogenous compounds in the atmosphere. These can be carried over long distances and deposited on glacier surfaces (Kozak et al, 2013) and have a significant impact upon nutrient cycling and possibly rates of activity and biomass production in these areas (although more research is required to confirm or deny this). Outside of the cryosphere, human activity has a well-known and marked effect upon ecosystem dynamics, particularly resulting from industrial emissions, road vehicles and the use of agricultural fertilisers.
Arctic/Antarctic
There is likely to be significant variation in nitrogen cycling in supraglacial habitats in the Arctic and Antarctic, largely due to the freezing of cryoconite hole ‘lids’ in Antarctica. These lids are formed when the surface of the melt water overlying cryoconite sediment in cryoconite holes freezes. This isolates the cryoconite hole from atmospheric exchanges and input from snowmelt. The process of nitrogen cycling in these pseudo-closed systems is uncertain, but is likely to be characterised by organic molecule recycling rather than fixation of dinitrogen, and may lead to extreme hydrochemical conditions (Tranter, 2004). There may also be significant differences between stable large glacier / ice sheet locations and alpine / ice-marginal zones where inputs and processes are more dynamic.
REF
Bayley, W.S. 1891. Mineralogy and Petrography. The American Naturalist, 25 (290): 138-146
Dittmar, T. , Fitznar, H. P. and Kattner, G. (2001): Origin and biogeochemical cycling of organic nitrogen in the eastern Arctic Ocean as evident from D- and L-amino acids , Geochimica et Cosmochimica Acta, 65 (22), pp. 4103-4114 .
Hodson A.: Biogeochemistry of snowmelt in an Antarctic glacial ecosystem, Water Resources Res., 42, doi: 10.1029/2005WR004311, 2006.
Hodson A.J., Mumford P.N., Kohler J. and Wynn P.M.: The High Arctic glacial ecosystem: New insights from nutrient budgets, Biogeochemistry, 72, 233-256, 2005.
Hodson A., Anesio A.M., Tranter M., Fountain A., Osborn M., Priscu J., Laybourn-Parry J. and Sattler B.: Glacial ecosystems, Ecological Monographs, 78, 41-67, doi:10.1890/07-0187.1, 2008.
Hodson A., Roberts T.J., Engvall A.-C., Holmen K. and Mumford P.: Glacier ecosystem response to episodic nitrogen enrichment in Svalbard, European High Arctic, Biogeochemistry, doi: 10.1007/s10533-009-9384-y, 2009.
Hodson, A., Roberts, T.J., Engvall, A-C., Holmen, K., Mumford, P. 2010. Glacier ecosystem response to episodic nitrogen enrichment in Svalbard, European High Arctic. Biogeosciences, 98 (1-3): 171-184
Jones, H.G. 1999. The ecology of snow-covered systems: a brief overview of nutrient cycling and life in the cold, Hydrological Processes, 13, 2135 – 2147.11
Kozak, K., Polkowska, Z., Ruman, M., Kozioł, K., Namieśnik, J. 2013. Analytical studies on the environmental state of the Svalbard archipelago provide a critical source of information about anthropogenic global impact. Trends in Analytical Chemistry, 50: 107 – 126
Nordin, A., Schmidt, I.K., Shaver, G.R. 2004.Nitrogen uptake by Arctic soil microbes and plants in relation to soil nitrogen supply. Ecology 85: 955–962
Olivier S., Blaser C., Brütsch S., Frolova N., Gäggeler H.W., Henderson K.A., Palmer A.S., Papina T., and Schwikowski M.: Temporal variations of mineral dust, biogenic tracers, and anthropogenic species during the past two centuries from Belukha ice core, Siberian Altai, J. Geophys. Res., 111, D05309, 2006.
Sawstrom, C., Karlsson, J., Laybourn-Parry, J., Graneli, W., (2009), Zooplankton feeding on algae and bacteria under ice in Lake Druzhby, East Antarctica. Polar Biology, 32(8), 1195-1202
Telling, J., A. M. Anesio, M. Tranter, T. Irvine-Fynn, A. Hodson, C. Butler, and J. Wadham (2011),Nitrogen fixation on Arctic glaciers, Svalbard, J. Geophys. Res., 116, G03039,
Telling, J., Stibal, M., Anesio, A.M., Tranter, M.L., Nias, I., Cook, J., Lis, G., Wadham, J.L., Sole, A., Nienow, P., Hodson, A.J. 2012. Microbial nitrogen cycling on the Greenland Ice Sheet. Biogeosciences,
Tockner, K., Malard, F., Uehlinger, U. and Ward, J. V. (2002) Nutrients and organic matter in a glacial river-floodplain system (Val Roseg, Switzerland). Limnology and Oceanography, 47, 266-277.
Tranter, M., FountainA., FritsenC., Lyons, B., Priscu, J., Statham, P., and Welch, K., 2004. Extreme hydrochemical conditions in natural microcosms entombed within Antarctic ice. Hydrological Processes, 18, 379-387.
Williams, M., Brooks, P.D., Mosier , M. 1996. Mineral nitrogen transformations in and under seasonal snow in a high-elevation catchment, Rocky Mountains, USA, Water Resources Research, 32, 3175-31856
Wynn P.M., Hodson A.J., Heaton T.H.E. and Chenery S.R.: Nitrate production beneath a High Arctic glacier, Svalbard, Chem. Geol. 244, 88-102, 2007

2 thoughts on “Nutrient Cycling on Glaciers 1: Nitrogen”