Carbon cycling on glaciers has received a lot of attention over the past decade because it impacts glacier albedo and therefore melt rates, as well as regional atmospheric carbon concentrations.
Atmospheric carbon concentrations and glacier retreat are known to be tightly coupled at a wide range of spatial and temporal scales. This article will concentrate upon relatively fast cycling through supraglacial microbial communities rather than tackling permafrost or millennial – geologic timescales. Following the structure of the N-cycling article, the fundamentals of global carbon cycling will be introduced, providing a background for exploring supraglacial carbon cycling afterwards.
The Carbon Cycle
Carbon is an absolutely crucial element fundamentally underpinning life on earth. It is the backbone of organic molecules from which life is composed; it is a primary constituent of the food we eat and the ground we inhabit.
Carbon continuously cycles between all components of the earth system, from the global scale over geologic time to the microscopic scale over fractions of seconds; between the hydrosphere, atmosphere, biosphere, cryosphere, lithosphere and anthrosphere. At the individual micro-organism scale, heterotrophs metabolise organic carbon to provide energy and release inorganic carbon into the atmosphere, while autotrophs use atmospheric carbon to synthesise organic molecules.
The constant flux of carbon molecules between various stores in the earth system is known as the carbon cycle. Human activities in contemporary times have disturbed the carbon budget – adding atmospheric carbon faster than the earth can absorb it into it’s other stores, and consequently temperatures have risen.
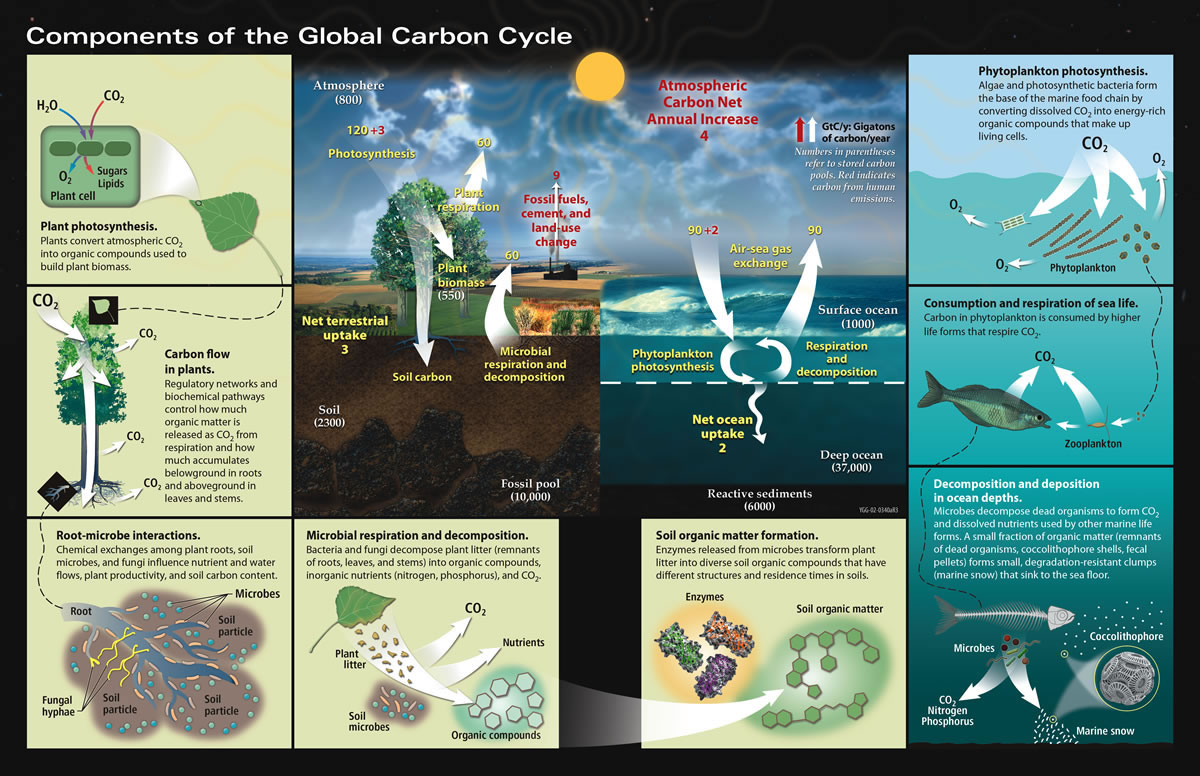
The diagram above shows the major components of the global carbon cycle. These components can be broadly split into two superimposed cycles: the slow C cycle and the fast C cycle:
Slow C Cycle:
The slow C cycle involves the incorporation of carbon into rocks. Atmospheric carbon reacts with water vapour to form carbonic acid, which reacts with silicate rocks to form bicarbonate ions. These ions find their way to the ocean via groundwater and rivers. In the ocean, carbon from bicarbonate ions is incorporated into the shells of marine organisms. When the organisms die, their shells collect on the seabed and are eventually compacted into carbonate rocks. Over geologic time, these are subducted into the mantle and released back into the atmosphere as CO2 via out-gassing during volcanic eruptions.
Fast C Cycle:
Superimposed upon the slow carbon cycle are much more rapid fluxes comprising the fast C cycle. These processes take days to decades and involve the transfer of carbon between the ocean, atmosphere, soils and organisms. Plants and phytoplankton absorb atmospheric CO2 and convert it – in the presence if sunlight – into sugars via a process called photosynthesis. This is called ‘fixing’, as atmospheric carbon is converted into larger molecules that have various uses within the biosphere. The first is as an energy source for the plants which fixed the atmospheric carbon in the first place (autotrophs). Secondly, they provide an energy source for organisms that eat the autotrophs (heterotrophs). Thirdly, plants die and decay. Finally, plants can be combusted by fire. Each of these four processes returns fixed carbon back into the atmosphere as carbon dioxide.
Carbon Cycling on Glaciers:
On glacier surfaces, carbon cycling is dominated by biotic processes. Although the supraglacial environment appears lifeless, it actually provides numerous habitats for microbial life – in particular algal blooms on the surface of the ice and small cylindrical depressions known as cryoconite holes (Yallop et al, 2012). Cryoconite holes are particularly active and biodiverse microbial habitats in which carbon is both fixed and respired (Hodson et al, 2010). Stibal et al (2012) described glacier surfaces as “factories that sequester organic carbon from the atmosphere and recycle recalcitrant organic carbon from various sources into more labile carbon substrates”, indicating that these highly active environments cycle carbon both produced in situ, predominantly via photosynthesis (aka autocthonous OC), and also carbon delivered from other sources (allochthonous OC).
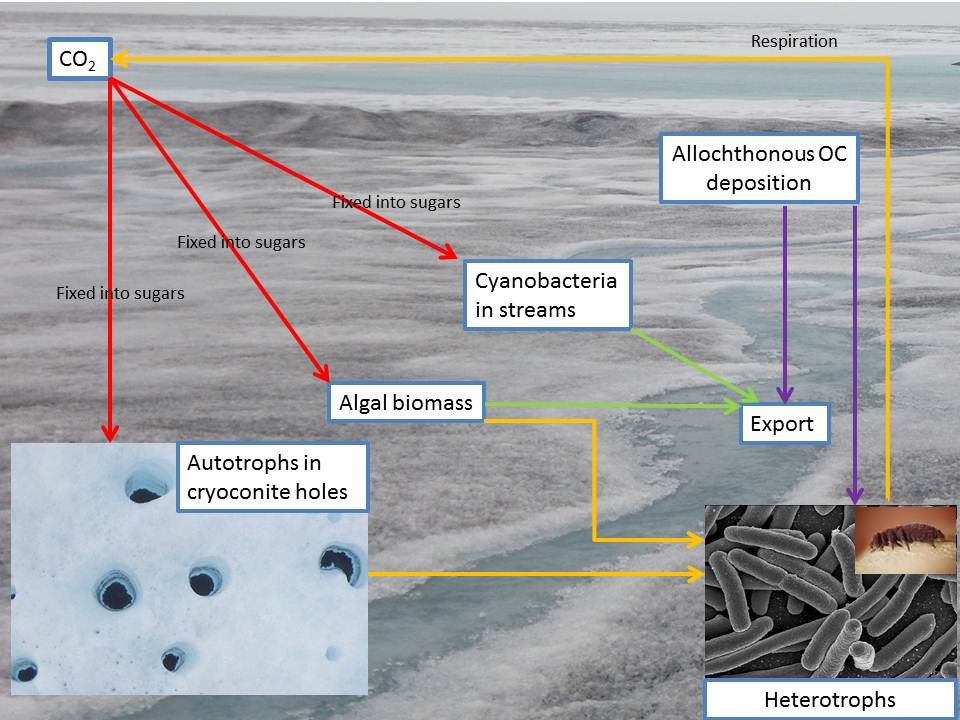
Atmospheric carbon is predominantly fixed by photoautotrophic microbes inhabiting glacier surfaces – mostly cyanobacteria and other algae. These organisms absorb carbon dioxide and fix it into sugars. In surface algal blooms these sugars are mostly used for microbial processes within the algae such as growth – adding biomass to the ice surface (Yallop et al, 2012). In cryoconite holes they are also used as a primary source of energy for higher organisms. When heterotrophic microbes feed upon primary producers some of the carbon which was used to build algal biomass is respired and released back into the atmosphere as CO2.
NEP
Net ecosystem productivity (NEP) describes the balance between autotrophy (fixing of CO2 into sugars) and heterotrophy (metabolism of sugars back into atmospheric CO2). This has been a particular focus for glacier microbiologists because not only does NEP determine whether a community represents a carbon sink or a carbon source (e.g. Stibal et al, 2008; Hodson et al, 2010; Cook et al, 2012), but also determines whether there is an overall increase in the amount of dark organic material on a glacier surface. The darker the surface, the more heat it absorbs and the faster it melts. Therefore, supraglacial carbon cycling might play a crucial role in the rate at which a glacier melts. Furthermore, autotrophic activity results in the production of dark humic material, photoprotective pigments and EPS (extracellular polymeris substances) (Langford et al, 2010) which actually give biomass a darker colour as well as just increasing its volume.
Allochthonous and Autochthonous Carbon
Microbes on glacier surfaces utilise carbon synthesised by autotrophs in situ (autochthonous), and also carbon deposited on the glacier from elsewhere (allochthonous). Autocthonous organic carbon is produced on ice surfaces as cellular biomass or cell exudates. Allochthonous organic carbon is deposited by wind – fragments of flora delivered from nearby deglaciated regions. Also black carbon from soot and anthropogenic pollutants are deposited from distant sources. Autochthonous and both types of allochthonous organic carbon provide energy for bacteria. The fraction of the total organic carbon originating from each of the above sources varies spatially and temporally, and has been the focus of much research (e.g. Stibal et al, 2008). It seems that the interiors of large ice sheets and glaciers are generally characterised by abundant autochthonous organic carbon providing substrate for community respiration – suggesting they represent carbon sinks – whereas ice sheet margins and small glaciers are likely dominated by local allochthonous carbon and act as net carbon sources (Stibal et al, 2012; Telling et al, 2012).
Export
Carbon fixed on glacier surfaces can also be redistributed. This is usually by entrainment into supraglacial streams or transport in solution through the low density ice that makes up the weathering crust (top ~2m of ice surface) (Irvine-Fynn and Edwards, 2013). Whole cells and organic carbon molecules dissolved in water (DOC) are transferred between habitats within the supraglacial zone and other glacial environments, as well as being redistributed to extra-glacial regions. This is likely to be important for seeding other environments (Wilhelm et al, 2013), such as proglacial streams, glacier fed lakes and the subglacial zone, as well as providing nutrients and energy sources for downstream ecosystems, and has recently been appreciated as an important factor in determining downstream biodiversity and community structure.
More information
For a bit more detail, see the article I wrote here and follow the references therein!
REF:
Cook, J.M., Hodson, A.J., Anesio, A.M., Hanna, E., Yallop, M., Stibal, M., Telling, J., Huybrechts, P. 2012. An improved estimate of microbially mediated carbon fluxes from the Greenland Ice Sheet. Journal of Glaciology, 58 (212)
Hodson, A., et al. 2010. A glacier respires: quantifying the istribution and and respiration CO2 flux of cryoconite across an entire Arctic supraglacial ecosystem. Journal of Geophysical Research, 112: G04S36
Irvine-Fynn, T. D. L. and Edwards, A. (2013), A frozen asset: The potential of flow cytometry in constraining the glacial biome. Cytometry. doi: 10.1002/cyto.a.22411
Langford, H., Hodson, A.J., Banwart, S., Boggild, C. 2010. The microstructure and biogeochemistry of Arctic cryoconite granules. Annals of Glaciology, 51 (56): 87-94
Stibal, M., Tranter, M., Benning, L., Rehak, J. 2008. Microbial primary production on glacier surfaces is insignificant in comparison with allochthonous organic carbon input. Environmental Microbiology, 10: 2172-2178
Stibal, M., Sabacka, M., Zarsky, J. 2012. Biological processes on glacier and ice sheet surfaces. Nature Geoscience, 5: 771-774
Telling, J. et al. 2012. Controls on the autochthonous production and respiration of organic matter in cryoconite holes on High Arctic glaciers. Journal of Geophysical Research, 117, G01017
Wilhelm, L., Singer, G.A., Fasching, C., Battin, T.J., Besemer, K. 2013. Microbial biodiversity in glacier-fed streams. ISME Journal, 7: 1651 – 1660
Yallop, M.L., Anesio, A.J., Perkins, R.G., Cook, J., Telling, J., Fagan, D., MacFarlane, J., Stibal, M., Barker, G., Bellas, C., Hodson, A., Tranter, M., Wadham, J., Roberts, N.W. 2012. Photophysiology and albedo-changing potential of the ice-algal community on the surface of the Greenland ice sheet. ISME Journal, 6: 2302 – 2313

One thought on “Nutrient Cycling on Glaciers 2: Carbon”