C Flux Modelling
To date, three attempts have been made to model carbon (C) fluxes in the supraglacial environment, all in the past five years. These models tried to reconcile ‘snapshot’ measurements of net ecosystem productivity (relative rates of photosynthesis and respiration – NEP) made at a small number of sites with atmospheric carbon fluxes at the local, regional and even global scale.
Why do we care?
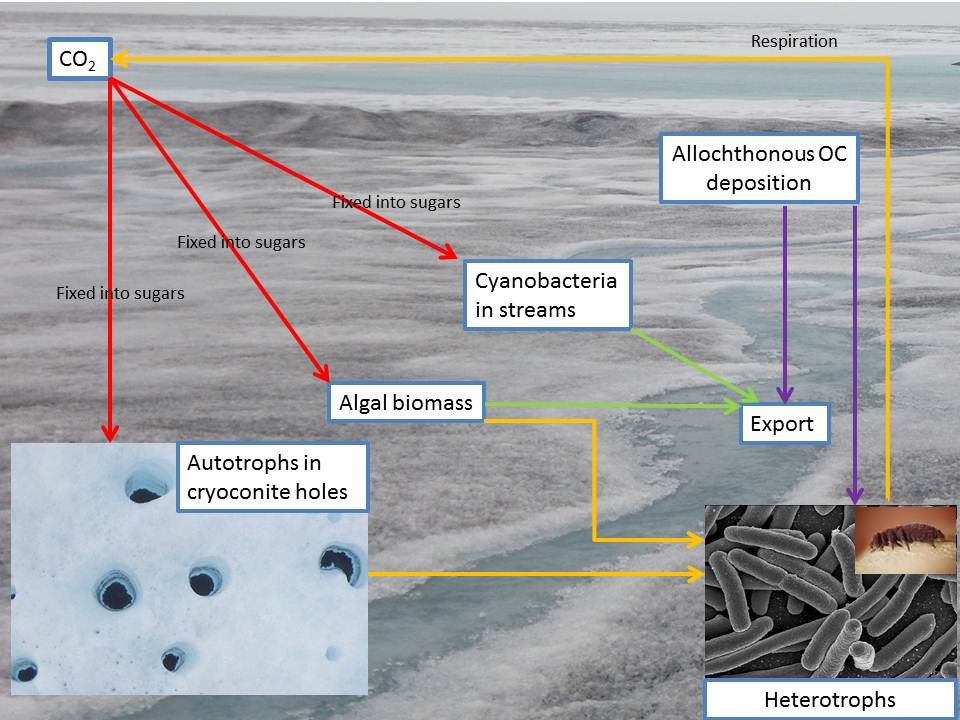
Nutrient fluxes on glaciers were largely overlooked until the past decade. However, fixation of CO2 by photosynthesis on glacier surfaces not only alters the concentration of this important greenhouse gas in the atmosphere, but also increases biomass on glacier surfaces (e.g. Hodson et al, 2008). This biomass is dark, potentially resides on glacier surfaces for long periods of time, and can therefore enhance glacier melt rates (Irvine-Fynn et al, 2011). This process is complicated by the opposing process of respiration, whereby fixed carbon is released back into the atmosphere as CO2. It is further complicated by non-biological processes (Stibal et al, 2012) such as meltwater runoff and seasonal snow deposition and melt. The net result of these factors defines not only the NEP, but also the abundance of various microorganisms that could eventually contribute genes, nutrients and cells to other ecosystems (e.g. Edwards et al, 2013). Modelling C fluxes in the supraglacial zone represents a first step in understanding the mechanisms and impacts of microbial activity in this environment.
The First Go:
Anesio et al (2009) made the first attempt to model supraglacial C fluxes at the global scale (excluding Antarctica). They averaged measurements of NEP from glacier surfaces in Svalbard, Greenland and the European Alps and then assumed them to be globally representative and temporally constant throughout a simulated melt season. The melt season was conservative in duration to account for a uniform ablation zone extent. This means that ice surfaces worldwide were assumed to simultaneously ‘switch on’ at the beginning of the summer and ‘switch off’ again at the end, rather than gradual exposure of bare ice by snowline retreat. A global net autotrophic C flux of 64Gg per year was estimated. Despite the major limitations outlined above, Anesio et al (2009) provided a crucial first estimate and paved the way for future studies.
The second go:
Hodson et al (2010) realised that upscaling snapshot measurements to the global scale was unjustifiable, and instead attempted to estimate C fluxes from the Greenland ice sheet. Ablation zone extent evolved throughout a melt season using a runoff-retention model (Janssen and Huybrechts, 2000). Within the ablation zone, photosynthesis occurred only on bare ice surfaces, whereas respiration occurred in both bare ice and slush. C fluxes were then estimated per unit area based upon measured NEP, cryoconite coverage and mass loading obtained from short transects near Kangerlussuaq (SW Greenland). A net autotrophic C flux of in the range 20 – 180 Gg per year was estimated.
The third go:
Despite Hodson et al’s (2010) model incorporating a temporally variable bare ice extent, it still assumed measurements obtained from a short transect to be representative across the entire ablation zone. Cook et al (2012) showed this to be erroneous, and in fact found the greatest rates of microbial production, surface coverage by cryoconite and mass loading to occur far beyond the maximum extent of Hodson et al’s (2010) transects. Cook et al (2012) therefore provided a model that included an evolving bare ice zone and also spatially variable rates of microbial activity, surface coverage and mass loading, based upon field measurements made across the entire ablation zone. Further, the previous models had assumed that cryoconites was the only supraglacial habitat that contributed to C cycling, whereas Cook et al (2012) recognised an additional algal ecosystem and built it into their model. Additionally, the size of the Greenland ice sheet makes upscaling from small transects in the SW corner to the entire ice sheet scale unrealistic, and therefore Cook et al (2012) only upscaled to a 1600 km2 area of the ice sheet surrounding their field transect, suggesting that to upscale further, additional transect data should be obtained.
C flux estimates that incorporated spatial variability in ice surface properties were four times greater than estimates based upon spatially uniform data. Furthermore, the inclusion of algal blooms increased the C flux by eleven times compared to cryoconite-only models. This highlighted the importance of obtaining field measurements from the entire ablation zone and considering the contribution made by all supraglacial habitats. The 1600km 2 area of the Greenland ice sheet was suggested to be in a stable state of net autotrophy. Estimates were made following 3 degrees of global warming, showing enhanced net autotrophy, however no attempt was made to consider any hydrological or albedo feedbacks that likely influence the system.

Where we stand:
Cook et al’s (2012) model represents the most sophisticated model to date, yet it is fundamentally limited in a number of ways. Firstly, there remains an assumption that there are only two discrete ecosystems that contribute to supraglacial C fluxes. Second, it assumes that microbial activity, spatial coverage and mass loading are constant throughout a melt season and that there is no redistribution of debris or biomas. Third, transect measurements are assumed to be representative across the entire 1600 km2 area. Fourth, it assumes no changes in albedo, hydrology or other abiotic factors occur during the melt season and makes no attempt to model any associated feedbacks. Therefore, there is much further work to be done to improve our estimates of supraglacial C fluxes and to obtain enough data to make estimates beyond the local scale.
New models using extensive measurements of biological activity, surface coverage, mass loading and hydrological and albedo feedbacks for the complete range of supraglacial habitats on ice surfaces worldwide are ideally needed, and they need to be built upon a deeper understanding of the complex processes operating on ice surfaces. Nevertheless, the three papers outlined above encourage an appreciation of the supraglacial zone as an important biome, and draw attention to its potential significance for global atmospheric exchanges.
REF:
Anesio, A.M; Hodson, A.j; Fritz, A; Psenner, R; Sattler, B (2009) ‘High microbial activity on glaciers: importance to the global carbon cycle’ Global Change Biology, Vol.15 (4): 955-960
Hodson, A.J; Anesio, A.M; Tranter, M; Fountain, A; Osborn, M; Priscu, J; Laybourn-Parry, J; Sattler, B (2008) ‘Glacial Ecosystems’ Ecological Monographs, Vol.78 (1): 41-67
Hodson, A.J., Bøggild, C.E., Hanna, E., Huybrechts, P., Langford, H., Cameron, K. and Houldsworth, A. (2010). The cryoconite ecosystem on the Greenland ice sheet. Annals of Glaciology, Vol.51 (56): 123-129.
Irvine-Fynn, T.D.L., Bridge, J., Hodson, A.J . 2011. In situ quantification of supraglacial cryoconite morphodynamics using time-lapse imaging: an example from Svalbard. Journal of Glaciology, 57 (204): 651 – 657
Jannsens, I., Huybrechts, P. 2000. The treatment of meltwater retardation in mass balance parameterizations of the Greenland Ice Sheet. Annals of Glaciology, 31: 133-140
Stibal, M., Telling, J., Cook, J., Man Mak, K., Hodson, A., Anesio, A. 2012. Environmental controls upon microbial abundance on the Greenland ice sheet: a multivariate analysis approach. Microbial Ecology, 63: 74-84
